Crop Adaptation: Weedy and Crop Wild Relatives as an Untapped Resource to Utilize Recent Increases in Atmospheric CO2

Mailman School of Public Health, Columbia University, New York, NY 10032, USA
Plants 2021, 10(1), 88; https://doi.org/10.3390/plants10010088
Submission received: 20 December 2020
/
Revised: 29 December 2020
/
Accepted: 31 December 2020
/
Published: 4 January 2021
(This article belongs to the Special Issue Crop Adaptation to Elevated CO2 and Temperature)
Abstract
:Adaptation measures are necessary to ensure the stability and performance of the food supply relative to anthropogenic climate change. Although a wide range of measures have been proposed (e.g., planting dates, crop choices, drought resistance), there may be a ubiquitous means to increase productivity relatively quickly. Numerous studies have shown that the projected increase in atmospheric CO2 can stimulate crop growth and seed yield with noted intra-specific differences within crop cultivars, suggesting potential differences to CO2 that could be exploited to enhance seed yield in the future. However, it is worth emphasizing that atmospheric CO2 has already risen substantially (≈27% since 1970) and that, at present, no active effort by breeders has been made to select for the CO2 increase that has already occurred. In contrast, for weedy or crop wild relatives (CWR), there are indications of evolutionary adaptation to these recent increases. While additional steps are needed, the identification and introgression of these CO2-sensitive traits into modern crop cultivars may be a simple and direct means to increase crop growth and seed yield.
1. Introduction
Maintaining food security is a seminal objective for the remainder of the century. While there are a number of recognized obstacles to achieve this objective, environmental limitations associated with unprecedented anthropogenic change threaten core aspects, including production, access, and quality. Specifically, climate change can alter abiotic environments, such as water availability (too much or too little), irregular temperatures, and extreme climatic events (flash droughts, derechos, etc.) [1,2]. Climatic-induced changes in biotic competition from agronomic pests (insects, disease, weeds) pose another significant constraint to global food production [3,4].
Such vulnerabilities within the agronomic food chain necessitate an immediate need to begin adapting crops to empirical threats associated with climatic change. Adaptation per se represents a wide range of approaches, but one facet of obvious consequence is genetics. In that regard, there are a number of exemplary and ongoing efforts to select for crop lines that can respond to climatic extremes, such as drought or extreme temperature [5,6,7].
There is another genetic approach that is becoming recognized as a potential adaptation tool: the selection of intra-specific variation in seed yield among C3 crop cultivars in response to projected increases in atmospheric CO2. Such an approach is being used to determine CO2 sensitivity in conjunction with economic yield for cassava [8], rice [9], soybean [10], wheat [11], inter alia. At present, there is ample evidence that considerable variation exists within crop cultivars for anticipated increases in CO2 and that selection for such variation holds promise as an adaptive means to increase crop yields.
Yet, the anticipated increase in atmospheric CO2 is, in the short term, slow, 2–3 ppm per year, and selection for cultivars for projected CO2 levels 30–50 years into the future does not address the current need to adapt crop systems.
On the other hand, atmospheric CO2 has already increased substantially from ≈325 to 412 ppm since 1970, which is an increase of ≈27%. Has this recent increase been exploited through ongoing artificial selection to choose current crop lines that are CO2 sensitive? Has the increase in CO2 been sufficient to begin evolutionary selection for increased growth and seed yield for weedy or crop wild relatives (CWR)? The objective of the current review is to compare and contrast selection efforts from breeders and nature with respect to CO2 sensitivity and potential seed yield for these two groups and to provide insight into metrics that could be of immediate (and future) benefit for utilizing CO2 to increase crop growth and seed yield.
2. Breeder Efforts to Select for CO2 Responsiveness in Crops
While intra-specific variation in response to future CO2 is being evidenced experimentally, there is no verification of any directed attempts by breeders to select for seed yield sensitivity to the increase in CO2 that has already occurred. It has been proposed that rapid-cycle breeding and new cultivar introductions could, over time, incrementally improve crop lines in adapting to new climates [12]. If true, then breeders could be already selecting (albeit passively) the most CO2-responsive cultivars over time. As such, there would be little need to initiate any active CO2 breeding programs to exploit recent (or projected) changes in CO2 with respect to seed yield.
Has such an approach worked to date? If modern, recently introduced lines are adapted to current CO2 levels (≈412 ppm), then they should demonstrate a greater CO2 response to recent CO2 increases relative to cultivars that were developed during the early twentieth century.
There are a number of studies that have explicitly tested this question. Sakai et al. [13] examined five japonica rice lines: ‘Aikoku’ (released in 1882), ‘Norin 8ʹ (1934), ‘Koshihikari’ (1956), ‘Akihikari’ (1976), and ‘Akidawara’ (2009). No differences in seed yield were noted between the oldest line (Aikoku, 1882) and the newest line (Akidawara, 2009) to increased CO2 (19 and 19.3%, increase at 600 ppm CO2, relative to ambient, respectively). For wheat, yield responses to rising CO2 actually declined with the release of newer cultivars [14,15]. For oat, Ziska and Blumenthal [16] examined the growth and vegetative characteristics of cultivated oat (Avena sativa L.) from seven geographical locations to CO2 concentration increases that corresponded roughly to the CO2 from the 1920s (300 ppm), as well as current (400 ppm) and mid-21st century projected levels (500 ppm). Newer lines were less responsive than older lines to rising CO2 in terms of both leaf area and tiller number. Overall, there is little evidence that cultivars in current use are those best adapted to maximize productivity in response to increasing atmospheric CO2 (but see [17] for barley).
Interestingly, for the oat study [16], significant age × CO2 interactions were observed with greater phenotypic variation noted for the older cultivars (Table 1). Diminished variance and increasing genetic uniformity is not unexpected, given increased farm size and greater mechanization during the 20th century [18]. However, uniformity can also limit responses to environmental factors, including rising CO2 [19], suggesting that the ability to respond to rising CO2 or other environmental perturbations may be constrained through modern breeding efforts. Overall, the narrow genetic base of modern cultivars may constitute a major bottleneck for crop improvement efforts [20,21,22].
3. Weedy and Wild Crop Relatives
Crop wild relatives (CWR) are those undomesticated “cousins” of cultivated crop lines that represent a potential untapped genetic resource that could be used as a means to adapt to new pests or abiotic changes, including, at least theoretically, increased levels of CO2. Of course, there are no directed efforts to adapt CWR to the recent increase in CO2; however, it is worth determining if any evolutionary adaptive changes that have allowed CWRs to adapt to recent CO2 increases have occurred.
It can be argued that such evolutionary changes are unlikely given the recent increase in CO2. However, the traditional paradigm of weed evolution as a very slow process is incomplete, and there are a number of examples demonstrating that rapid evolutionary change (years or decades) can occur within weed biology, (e.g., Microstegium vimineum, [23]; Lythrum salicaria, [24]; Brassica rapa, [25]; Avena fatua, [26]). In turn, such changes could include evolution in response to anthropogenic climate change [27,28,29].
At present, there is initial evidence indicating that recent increases in CO2 may have already altered the adaptive response of some annual weeds. For example, Bunce [30] examined recent increases in CO2 on the growth response of four annual weeds over a narrow CO2 range (90 ppm below and above ambient) and demonstrated that the efficiency by which these weeds utilized CO2 declined at concentrations above ambient, indicating that these weeds had adapted to recent CO2 increases.
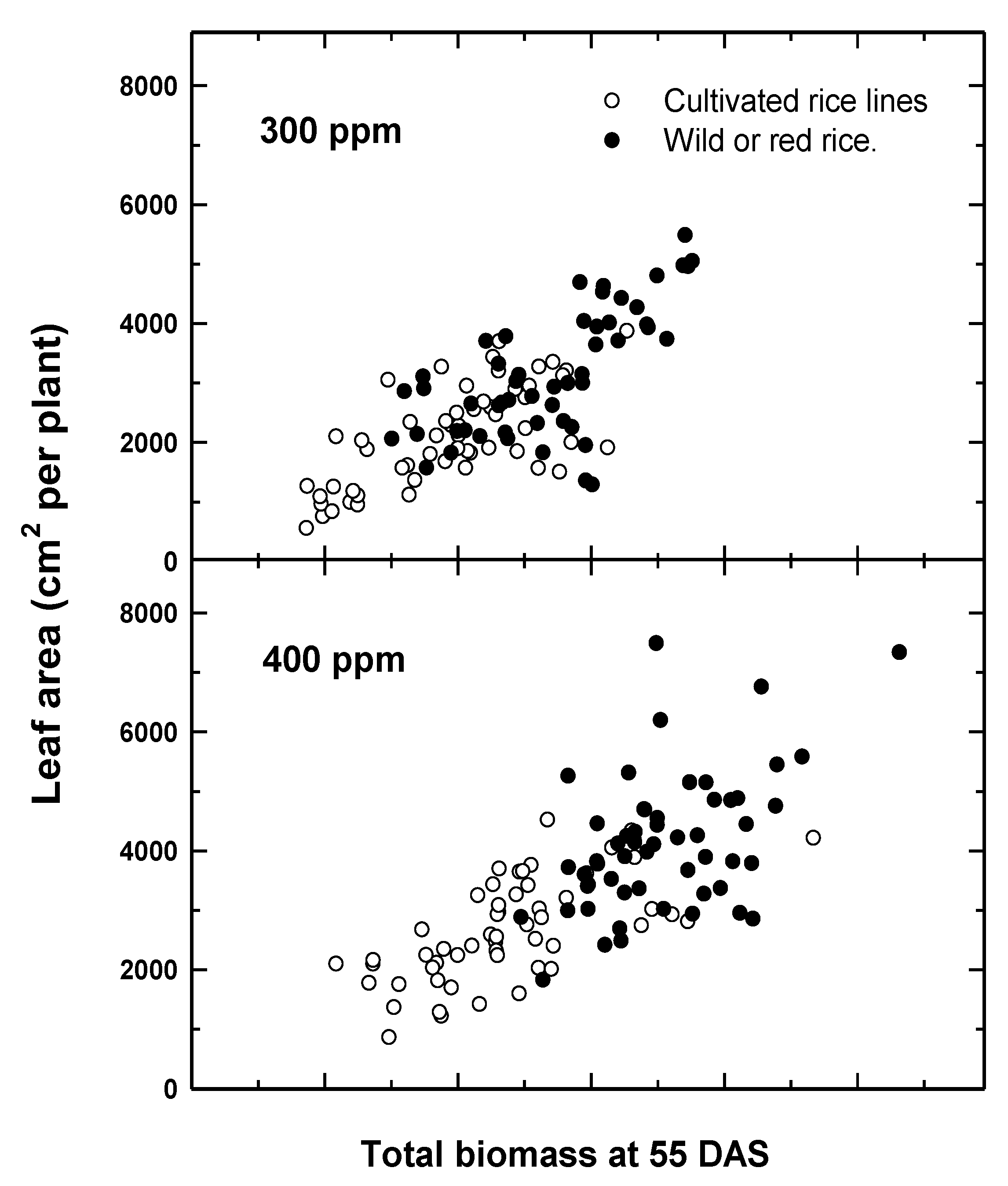
With respect to CWR, comparisons of relative fitness to cultivated lines suggest differential adaptation to recent CO2 increases. Comparisons of six cultivated and six wild or weedy biotypes of rice indicated a greater overall growth response among wild relative to cultivated rice to recent (300–400 ppm) increases in CO2 [31] (Figure 1), suggesting that the rapid evolution of weedy biotypes may have increased their fitness relative to the crop. Greater seed yields were also recorded for Stuttgart, a CWR to rice, compared to Clearfield, a cultivated rice line, for the same recent CO2 increase [32]. Similarly, using a resurrection approach [33], seeds of two temporally distinct populations of CWR wild oat (Avena fatua L.) from the same location, one from the 1960s and one from 2014, (a relative CO2 increase of 80 ppm, or 25% from 1960) demonstrated different competitive abilities against a cultivated oat (A. sativa) line, with the more recent (2014) A. fatua population having greater growth and competitive ability at current CO2 levels [34].
4. Differences in Selection
Additional data to confirm and expand upon these results is obviously needed. However, it is important to recognize differences within the selection process for cultivated and CWR biotypes. Weeds and wild crop relatives have been subject to natural environmental perturbations for millennia and have as a consequence maintained a much higher level of genetic diversity [21]. Both are characterized by rapid growth, high seed production, environmental plasticity, and genetic variability, and they are considered highly adaptable [35,36]. Such plant species, with short generation times and high seed production, generally show more rapid rates of molecular evolution [37].
Conversely, it is generally recognized that advances in plant breeding are associated with recurrent selection, usually in field environments. As a consequence, selection for say, pest resistance, should be occurring concurrently with rising CO2, and, as a result, reflect CO2 adaptation. However, plant breeding is a long-term process that can extend over decades, and indirect selection for yield under field conditions is likely to be inefficient because yield is related to a number of abiotic and biotic factors. The unintended consequences of recurring selection are that genetic variation and associated phenotypes can be reduced relative to the available hereditary potential.
Genetic exchange among breeders has also shifted over time. As documented by Atlin [12], prior to the 1990s, breeders typically exchanged varieties. However, as Genetically modified organism (GMO) lines were introduced and breeding became more commercialized, Intellectual Property (IP) protection increased, resulting in the current U.S. practice of issuing utility patents that prevent proprietary lines from being used as parents by other breeders. Unfortunately, simultaneous with these changes, a number of countries recognized that their own indigenous crops were unique genetic resources and restricted their usage in public breeding programs. Consequently, obtaining elite or novel varieties internationally has become restrictive. Overall, such actions have reduced reliance on publically available seed sources, reducing genetic diversity [38].
Given the need for the mechanization of large land holdings and economic consistency in response to water and fertilizer so as to achieve high crop productivity, dedomestication and genetic uniformity are necessary, even if such practices lead to “Domestication Bottlenecks” [39]. However, such uniformity in management may also limit the extent of genetic variation in response to environmental changes, such as CO2 [40].
Overall, opportunities for increasing production may be missed if we assume that current breeding efforts have resulted in crop plants that are adapted to the recent increase in CO2 concentration. Rather, it suggests that CWR and natural selection may serve as a starting point for active intervention to enhance genetic diversity to meet new environmental uncertainty and optimize crop yields to ongoing CO2 increases. In addition, land races, populations of a cultivated plant that are distinct to a given geographic and environmental locale, are worth additional evaluation and may also provide a useful genetic resource in that regard.
5. Challenges and Next Steps
If CWR represent an opportunity to exploit the recent increase in CO2, it is an opportunity that includes a number of pragmatic challenges. What follows is by no means inclusive but rather representative of next steps.
5.1. Phenotypic and Genotypic CO2 Sensitivity Traits
Greater effort is necessary to document and determine those traits that may have already contributed to a greater sensitivity to recent CO2 increases (e.g., red rice). A consensus is needed to identify those phenological, morphological, and/or physiological characteristics that are associated with CO2 responsiveness. Initial studies have suggested different organismal levels associated with CO2 sensitivity including genetic (e.g., carbohydrate regulation of RNA, [41], biochemical (e.g., Rubisco activase) [42], leaf (e.g., stomatal density [43] or photosynthesis [44], whole plant (relative growth rate) [45], management (e.g., planting density) [46] and canopy (e.g., nitrogen applications) [47], but specific organismal characteristics consistently associated with CO2 responsiveness and crop yield for CWRs have not been identified. A definitive set of these parameters is necessary for breeders to select CO2-sensitive crop archetypes.
5.2. Introgression of CWR Traits
While CWRs can make an effective contribution to broadening the genetic diversity of crops, their direct use in breeding has primarily focused on introgressing loci for disease resistance, not abiotic stress [48,49]. This is due, in part, to the presence of objectionable traits in CWRs (e.g., transfer of undesirable QTLs) as well as breeding barriers with the crop.
Initiatives have commenced that aim to adapt agricultural sustainability to climate change through the use of CWRs to broaden and improve the cultivated gene pool as part of the International Treaty on Plant Genetic Resources for Food and Agriculture (ITPGRFA, [50]). This effort is designed to collect, preserve, and prepare CWRs for evaluation and potential adaptation of crops to climate change. Other initiatives, such as Diversity Seek (DivSeek) are underway to begin evaluating the potential of crop and wild relative diversity present within gene banks [51].
To overcome difficulties in facilitating gene introgression, Prohens et al. [52] have suggested a novel approach, ‘introgressiomics’, a mass-scale expansion of plant materials and populations that confer genetic introgressions from CWRs into crops. Their goal is to generate chromosome substitution lines (CSLs), introgression lines (ILs), and multi-parent advanced inter-cross (MAGIC) populations through the use of marker-assisted selection as a means to characterize genetic traits present in CWRs, but to also develop genetically relevant elite materials that can be incorporated into breeding programs as a means to adapt crops to climatic change.
5.3. Additional Climatic Variables
The climatic consequences of increasing CO2 are obvious and include increases in surface temperature, changes in precipitation and extreme events that will have negative consequence for crop productivity. Temperatures, especially during anthesis, may be critical in maintaining yield performance [53]. In addition, there are numerous studies indicating that rising temperature per se may negate any stimulatory effect for projected CO2 increases [54,55,56].
At present, evaluations of CWR to both recent increases in CO2 and temperature are unavailable; however, it is of interest to note one seminal study that has examined Indica, Indica-like, Japonica, and CWR of rice to projected CO2 concentrations four temperature treatments [57]. They reported an increased yield sensitivity to high temperature stress at higher (600 ppm) CO2, but their results also showed that CWR for rice demonstrated superior CO2 x temperature interactions with respect to yield, supporting the idea of using wild or unadapted gene pools in rice to enhance breeding efforts for climate change adaptation.
In any case, a fundamental challenge in CO2 selection will be to consider multiple environmental interactions with a merited focus on temperature and moisture conditions to assess possible negative interactions with respect to yield. By necessity, such selections will include evaluations of multiple-gene responses.
5.4. Nutritional Considerations
For projected CO2 increases, there is considerable evidence indicating that stimulatory effects of increased CO2 may be accompanied by declines in nutritional quality, including but not limited to protein and minerals [58,59,60,61]. The basis for the CO2-induced changes in crop quality are still being elucidated, in part because there are a number of biological and physical processes that are influenced by increasing CO2 [62]. However, any efforts to adapt CWRs and cultivars to recent CO2 increases must include concurrent selection efforts or the co-development of suitable management practices that will maintain the desired quality and nutritional characteristics necessary for human health.
5.5. Scaling Up to the Field Level
Genomic and molecular traits that are assimilated into new cultivars may increase performance at the whole plant or leaf level; however, as emphasized by Sinclair et al. [63], such improvements need to scale up to field responses. As such, the identification of CO2-induced increases in the photosynthetic rate or seed yield of single leaves or whole plants in the laboratory require complementary management approaches that will illicit similar responses in the field. In this regard, it may be worthwhile to examine historical trends by cultivar. For example, comparisons of yield performance from 1950s cultivars, relative to the same cultivars under today’s CO2 concentration using similar metrics (row spacing, soil types, pesticide usage) could help identify cultivar x CO2 sensitivity and, potentially, elucidate management practices that could maximize CO2 yield responses in situ.
6. Conclusions
The challenge of adapting to an uncertain climate is paramount to maintaining global food security. High yielding crop varieties with tolerance to biotic and abiotic stresses associated with climatic change are needed to meet this challenge. Currently, there is recognition that the current genetic base of modern cultivars may be too narrow for crop improvement efforts in that regard. In turn, this may reflect a lack of genetic ideotypes that encompass traits associated with climatic change or uncertainty associated with the rate of change. As such, the use of CWR (or landraces) may be a means of enhancing genetic diversity of cultivated crops as a means to adapt to these changes.
Among adaptation efforts, there is an opportunity to exploit the recent increase in atmospheric CO2 as a means to stimulate plant growth and yield. Currently, comparisons of cultivated and CWR indicate differential selection in regard to this change, with initial evidence suggesting evolutionary adaptation for some CWR, e.g., red rice. Introgression of these traits into current cultivars, and appropriate management practices, may provide a means to utilize the increase in CO2 that has already occurred (≈27% increase since 1970) to stimulate seed yield.
At present, there is no empirical evidence that breeders are selecting for CO2 responsiveness. This may be due, in part, to current economic and agronomic practices associated with modern agriculture. Yet, it seems surprising that CO2 as a potential adaptation strategy to maintain global food production is not being utilized. One can ask if other abiotic resources such as sunlight, water, or nutrients had increased to a similar extent in recent decades whether incentives to optimize that increase would be underway.
Adaptation to recent CO2 increases will not be a complete solution to the complications associated with climate change. However, adaptation may represent one of the simplest research strategies to help maintain global food security relative to the anthropogenic stresses associated with climatic change. As such, it is hoped that this review can serve as a sounding board and starting point for additional efforts.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The article is a review synopsis and does not contain original data.
Acknowledgments
The author thanks James Bunce for his invitation and for his years of mentorship.
Conflicts of Interest
The author declares no conflict of interest.
References
- Harrison, M.T.; Cullen, B.R.; Rawnsley, R.P. Modelling the sensitivity of agricultural systems to climate change and extreme climatic events. Agric. Syst. 2016, 148, 135–148. [Google Scholar] [CrossRef]
- Petrie, M.D.; Bradford, J.B.; Lauenroth, W.K.; Schlaepfer, D.R.; Andrews, C.M.; Bell, D.M. Non-analog increases to air, surface, and belowground temperature extreme events due to climate change. Clim. Chang. 2020, 158, 1–24. [Google Scholar]
- Deutsch, C.A.; Tewksbury, J.J.; Tigchelaar, M.; Battisti, D.S.; Merrill, S.C.; Huey, R.B.; Naylor, R.L. Increase in crop losses to insect pests in a warming climate. Science 2018, 361, 916–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziska, L.H.; Bradley, B.A.; Wallace, R.D.; Bargeron, C.T.; LaForest, J.H.; Choudhury, R.A.; Garrett, K.A.; Vega, F.E. Climate change, carbon dioxide, and pest biology, managing the future: Coffee as a case study. Agronomy 2018, 8, 152. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Singh, G.S. Traditional agriculture: A climate-smart approach for sustainable food production. Energy Ecol. Environ. 2017, 2, 296–316. [Google Scholar] [CrossRef]
- Henry, R.J. Innovations in plant genetics adapting agriculture to climate change. Curr. Opin. Plant Biol. 2020, 56, 168–173. [Google Scholar] [CrossRef]
- Javadinejad, S.; Dara, R.; Jafary, F. Analysis and prioritization the effective factors on increasing farmers resilience under climate change and drought. Agric. Res. 2020, 9, 1–17. [Google Scholar] [CrossRef]
- Ruiz-Vera, U.M.; De Souza, A.P.; Ament, M.R.; Gleadow, R.M.; Ort, D.R. High sink strength prevents photosynthetic down-regulation in cassava grown at elevated CO2 concentration. J. Exp. Bot. 2020. [Google Scholar] [CrossRef]
- Hasegawa, T.; Sakai, H.; Tokida, T.; Nakamura, H.; Zhu, C.; Usui, Y.; Yoshimoto, M.; Fukuoka, M.; Wakatsuki, H.; Katayanagi, N.; et al. Rice cultivar responses to elevated CO2 at two free-air CO2 enrichment (FACE) sites in Japan. Funct. Plant Biol. 2012, 40, 148–159. [Google Scholar] [CrossRef] [Green Version]
- Bishop, K.A.; Betzelberger, A.M.; Long, S.P.; Ainsworth, E.A. Is there potential to adapt soybean (G lycine max M err.) to future [CO2]? An analysis of the yield response of 18 genotypes in free-air CO2 enrichment. Plant Cell Environ. 2015, 38, 1765–1774. [Google Scholar] [CrossRef]
- Bunce, J. Using FACE systems to screen wheat cultivars for yield increases at elevated CO2. Agronomy 2017, 7, 20. [Google Scholar] [CrossRef] [Green Version]
- Atlin, G.N.; Cairns, J.E.; Das, B. Rapid breeding and varietal replacement are critical to adaptation of cropping systems in the developing world to climate change. Glob. Food Sec. 2017, 12, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Sakai, H.; Tokida, T.; Usui, Y.; Nakamura, H.; Hasegawa, T. Yield responses to elevated CO2 concentration among Japanese rice cultivars released since 1882. Plant Prod. Sci. 2019, 22, 352–366. [Google Scholar] [CrossRef] [Green Version]
- Manderscheid, R.; Weigel, H.J. Photosynthetic and growth responses of old and modern spring wheat cultivars to atmospheric CO2 enrichment. Ag. Ecosys. Environ. 1997, 64, 65–73. [Google Scholar] [CrossRef]
- Ziska, L.H.; Morris, C.F.; Goins, E.W. Quantitative and qualitative evaluation of selected wheat varieties released since 1903 to increasing atmospheric carbon dioxide: Can yield sensitivity to carbon dioxide be a factor in wheat performance? Glob. Chang. Biol. 2004, 10, 1810–1819. [Google Scholar] [CrossRef]
- Ziska, L.H.; Blumenthal, D.M. Empirical selection of cultivated oat in response to rising atmospheric carbon dioxide. Crop Sci. 2007, 47, 1547–1552. [Google Scholar] [CrossRef] [Green Version]
- Schmid, I.; Franzaring, J.; Müller, M.; Brohon, N.; Calvo, O.C.; Högy, P.; Fangmeier, A. Effects of CO2 enrichment and drought on photosynthesis, growth and yield of an old and a modern barley cultivar. J. Agron. Crop Sci. 2016, 202, 81–95. [Google Scholar] [CrossRef]
- Doyle, J. Altered Harvest: Agriculture, Genetics and the Fate of the World’s Food Supply; Viking Press: New York, NY, USA, 1985. [Google Scholar]
- Reich, P.B.; Knops, J.; Tilman, D.; Craine, J.; Ellsworth, D.; Tjoelker, M.; Lee, T.; Wedin, D.; Naeem, S.; Bahauddin, D.; et al. Plant diversity enhances ecosystem responses to elevated CO2 and nitrogen deposition. Nature 2001, 410, 809–810. [Google Scholar] [CrossRef]
- Mammadov, J.; Buyyarapu, R.; Guttikonda, S.K.; Parliament, K.; Abdurakhmonov, I.Y.; Kumpatla, S.P. Wild relatives of maize, rice, cotton, and soybean: Treasure troves for tolerance to biotic and abiotic stresses. Front. Plant Sci. 2018, 9, 886. [Google Scholar] [CrossRef]
- Zhang, H.; Mittal, N.; Leamy, L.J.; Barazani, O.; Song, B.H. Back into the wild—Apply untapped genetic diversity of wild relatives for crop improvement. Evol. App. 2017, 10, 5–24. [Google Scholar] [CrossRef]
- Zhang, F.; Batley, J. Exploring the application of wild species for crop improvement in a changing climate. Curr. Opin. Plant Biol. 2020, 56, 218–222. [Google Scholar] [CrossRef] [PubMed]
- Novy, A.; Flory, S.L.; Hartman, J.M. Evidence for rapid evolution of phenology in an invasive grass. J. Evol. Biol. 2013, 26, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Colautti, R.I.; Barrett, S.C. Rapid adaptation to climate facilitates range expansion of an invasive plant. Science 2013, 342, 364–366. [Google Scholar] [CrossRef] [PubMed]
- Franks, S.J.; Sim, S.; Weis, A.E. Rapid evolution of flowering time by annual plant in response to a climate fluctuation. Proc. Natl. Acad. Sci. USA 2007, 104, 1278–1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naylor, J.M.; Jana, S. Genetic adaptation for seed dormancy in Avena fatua. Can. J. Bot. 1976, 54, 306–312. [Google Scholar] [CrossRef]
- Clements, D.R.; DiTommaso, A. Climate change and weed adaptation: Can evolution of invasive plants lead to greater range expansion than forecasted? Weed Res. 2011, 51, 227–240. [Google Scholar] [CrossRef]
- Ravet, K.; Patterson, E.L.; Krähmer, H.; Hamouzová, K.; Fan, L.; Jasieniuk, M.; Lawton-Rauh, A.; Malone, J.M.; McElroy, J.S.; Merotto, A., Jr.; et al. The power and potential of genomics in weed biology and management. Pest Manag. Sci. 2018, 74, 2216–2225. [Google Scholar] [CrossRef] [PubMed]
- Ziska, L.H.; Blumenthal, D.M.; Franks, S.J. Understanding the nexus of rising CO2, climate change, and evolution in weed biology. Invasive Plant Sci. Manag. 2019, 12, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Bunce, J.A. Are annual plants adapted to the current atmospheric concentration of carbon dioxide? Int. J. Plant Sci. 2001, 162, 1261–1266. [Google Scholar] [CrossRef]
- Ziska, L.H.; McClung, A. Differential response of cultivated and weedy (red) rice to recent and projected increases in atmospheric carbon dioxide. Agron. J. 2008, 100, 1259–1263. [Google Scholar] [CrossRef]
- Ziska, L.H.; Tomecek, M.B.; Gealy, D.R. Competitive interactions between cultivated and red rice as a function of recent and projected increases in atmospheric carbon dioxide. Agron. J. 2010, 102, 118–123. [Google Scholar] [CrossRef] [Green Version]
- Franks, S.J.; Hamann, E.; Weis, A.E. Using the resurrection approach to understand contemporary evolution in changing environments. Evol. App. 2018, 11, 17–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziska, L.H. Could recent increases in atmospheric CO2 have acted as a selection factor in Avena fatua populations? A case study of cultivated and wild oat competition. Weed Res. 2017, 57, 399–405. [Google Scholar] [CrossRef]
- Baker, H.G. The evolution of weeds. Ann. Rev. Ecol. Sys. 1974, 5, 1–24. [Google Scholar] [CrossRef]
- Radosevich, S.R.; Holt, J.S.; Ghersa, C. Weed Ecology: Implications for Management; John Wiley & Sons: New York, NY, USA, 1997. [Google Scholar]
- Smith, S.A.; Donoghue, M.J. Rates of molecular evolution are linked to life history in flowering plants. Science 2008, 322, 86–89. [Google Scholar] [CrossRef] [Green Version]
- Jacobsen, S.E.; Sørensen, M.; Pedersen, S.M.; Weiner, J. Feeding the world: Genetically modified crops versus agricultural biodiversity. Agron. Sust. Develop. 2013, 33, 651–662. [Google Scholar] [CrossRef] [Green Version]
- Tanksley, S.D.; McCouch, S.R. Seed banks and molecular maps: Unlocking genetic potential from the wild. Science 1997, 277, 1063–1066. [Google Scholar] [CrossRef] [Green Version]
- Treharne, K. The implications of the ‘greenhouse effect’ for fertilizers and agrochemicals. In The Greenhouse Effect and UK Agriculture; Bennet, R.C., Ed.; Ministry of Agriculture, Fisheries and Food: London, UK, 1989; pp. 67–78. [Google Scholar]
- Sheen, J. Feedback control of gene expression. Photosyn. Res. 1994, 39, 427–438. [Google Scholar] [CrossRef]
- Sage, R.F.; Coleman, J.R. Effects of low atmospheric CO2 on plants: More than a thing of the past. Trends Plant Sci. 2001, 6, 18–24. [Google Scholar] [CrossRef]
- Ward, J.K.; Kelly, J.K. Scaling up evolutionary responses to elevated CO2: Lessons from Arabidopsis. Ecol. Lett. 2004, 7, 427–440. [Google Scholar] [CrossRef]
- Bunce, J.A. Contrasting responses of seed yield to elevated carbon dioxide under field conditions within Phaseolus vulgaris. Agric. Ecosyst. Environ. 2008, 128, 219–234. [Google Scholar] [CrossRef]
- Poorter, H. Do slow-growing species and nutrientstressed plants respond relatively strongly to elevated CO2? Glob. Change Biol. 1998, 4, 693–697. [Google Scholar] [CrossRef]
- Shimono, H. Rice genotypes that respond strongly to elevated CO2, also respond strongly to low planting density. Agric. Ecosyst. Environ. 2011, 141, 240–243. [Google Scholar] [CrossRef]
- Ziska, L.H.; Weerakoon, W.; Namuco, O.S.; Pamplona, R. The influence of nitrogen on the elevated CO2 response in field-grown rice. Austral. J. Plant Physiol. 1996, 23, 45–52. [Google Scholar] [CrossRef]
- Hajjar, R.; Hodgkin, T. The use of wild relatives in crop improvement: A survey of developments over the last 20 years. Euphytica 2007, 156, 1–13. [Google Scholar] [CrossRef]
- Maxted, N.; Kell, S.P. Establishment of a global network for the in situ conservation of crop wild relatives: Status and needs. FAO Comm. Genet. Resour. Food Agric. Rome Italy 2009, 266, 3–94. [Google Scholar]
- Dempewolf, H.; Eastwood, R.J.; Guarino, L.; Khoury, C.K.; Müller, J.V.; Toll, J. Adapting agriculture to climate change: A global initiative to collect, conserve, and use crop wild relatives. Agroecol. Sustain. Food Syst. 2014, 38, 369–377. [Google Scholar] [CrossRef] [Green Version]
- Meyer, R.S. Encouraging metadata curation in the Diversity Seek initiative. Nat. Plants 2015, 1, 1–2. [Google Scholar] [CrossRef]
- Prohens, J.; Gramazio, P.; Plazas, M.; Dempewolf, H.; Kilian, B.; Díez, M.J.; Fita, A.; Herraiz, F.J.; Rodríguez-Burruezo, A.; Soler, S.; et al. Introgressiomics: A new approach for using crop wild relatives in breeding for adaptation to climate change. Euphytica 2017, 213, 158–166. [Google Scholar] [CrossRef]
- Schlenker, W.; Roberts, M.J. Nonlinear temperature effects indicate severe damages to US crop yields under climate change. Proc. Natl. Acad. Sci. USA 2009, 106, 15594–15598. [Google Scholar] [CrossRef] [Green Version]
- Chavan, S.G.; Duursma, R.A.; Tausz, M.; Ghannoum, O. Elevated CO2 alleviates the negative impact of heat stress on wheat physiology but not on grain yield. J. Exp. Bot. 2019, 70, 6447–6459. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Cai, C.; He, J.; Gu, J.; Zhu, G.; Zhang, W.; Zhu, J.; Liu, G. Yield, dry matter distribution and photosynthetic characteristics of rice under elevated CO2 and increased temperature conditions. Field Crops Res. 2020, 248, 107605. [Google Scholar] [CrossRef]
- Wang, J.; Wang, C.; Chen, N.; Xiong, Z.; Wolfe, D.; Zou, J. Response of rice production to elevated [CO2] and its interaction with rising temperature or nitrogen supply: A meta-analysis. Clim. Chang. 2015, 130, 529–543. [Google Scholar] [CrossRef]
- Wang, D.R.; Bunce, J.A.; Tomecek, M.B.; Gealy, D.; McClung, A.; McCouch, S.R.; Ziska, L.H. Evidence for divergence of response in Indica, Japonica, and wild rice to high CO 2× temperature interaction. Glob. Chang. Biol. 2016, 22, 2620–2632. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Ineson, P.; Scott, A. Elevated CO2 reduces the nitrogen concentration of plant tissues. Glob. Chang. Biol. 1998, 4, 43–54. [Google Scholar] [CrossRef]
- Loladze, I. Hidden shift of the ionome of plants exposed to elevated CO2 depletes minerals at the base of human nutrition. Elife 2014, 3, e02245. [Google Scholar] [CrossRef]
- Loladze, I. Rising atmospheric CO2 and human nutrition: Toward globally imbalanced plant stoichiometry? Trends Ecol. Evol. 2002, 17, 457–461. [Google Scholar] [CrossRef]
- Myers, S.S.; Zanobetti, A.; Kloog, I.; Huybers, P.; Leakey, A.D.; Bloom, A.J.; Carlisle, E.; Dietterich, L.H.; Fitzgerald, G.; Hasegawa, T.; et al. Increasing CO2 threatens human nutrition. Nature 2014, 510, 139–142. [Google Scholar] [CrossRef]
- McGrath, J.M.; Lobell, D.B. Reduction of transpiration and altered nutrient allocation contribute to nutrient decline of crops grown in elevated CO2 concentrations. Plant Cell Environ. 2013, 36, 697–705. [Google Scholar] [CrossRef]
- Sinclair, T.R.; Purcell, L.C.; Sneller, C.H. Crop transformation and the challenge to increase yield potential. Trends Plant Sci. 2004, 9, 70–75. [Google Scholar] [CrossRef]
Figure 1.
Response of six red rice (RR, filled circles) and six cultivated rice (CR, open circles) varieties to the recent change in atmospheric CO2 concentrations (300–400 ppm) for leaf area as a function of total above-ground biomass 55 days after sowing. Note the increase in growth for RR for the recent CO2 increase. Data are adapted from Ziska and McClung [31].
Figure 1.
Response of six red rice (RR, filled circles) and six cultivated rice (CR, open circles) varieties to the recent change in atmospheric CO2 concentrations (300–400 ppm) for leaf area as a function of total above-ground biomass 55 days after sowing. Note the increase in growth for RR for the recent CO2 increase. Data are adapted from Ziska and McClung [31].


{kind=link}
Table 1.
Significance of equality of variance for “old” and “new” oat cultivars for select vegetative and growth characteristics averaged over all CO2 concentrations. An asterisk indicates a greater degree of phenotypic variation. Note the greater degree of phenotypic variation for oat cultivars released in the 1920s. Data are from 16.
Table 1.
Significance of equality of variance for “old” and “new” oat cultivars for select vegetative and growth characteristics averaged over all CO2 concentrations. An asterisk indicates a greater degree of phenotypic variation. Note the greater degree of phenotypic variation for oat cultivars released in the 1920s. Data are from 16.
| Variable | “Old” | “New” |
|---|---|---|
| Leaf area | * | |
| Leaf wt. | ** | |
| Tiller wt. | ** | |
| Weight tiller−1 | *** | |
| Tiller No. | * | |
| Root wt. | * | |
| Total wt. | *** | |
| RGR | ** |
* p < 0.05; ** p < 0.01; *** p < 0.001. n.s. = not significant.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ziska, L.H. Crop Adaptation: Weedy and Crop Wild Relatives as an Untapped Resource to Utilize Recent Increases in Atmospheric CO2. Plants 2021, 10, 88. https://doi.org/10.3390/plants10010088
AMA Style
Ziska LH. Crop Adaptation: Weedy and Crop Wild Relatives as an Untapped Resource to Utilize Recent Increases in Atmospheric CO2. Plants. 2021; 10(1):88. https://doi.org/10.3390/plants10010088
Chicago/Turabian StyleZiska, Lewis H. 2021. "Crop Adaptation: Weedy and Crop Wild Relatives as an Untapped Resource to Utilize Recent Increases in Atmospheric CO2" Plants 10, no. 1: 88. https://doi.org/10.3390/plants10010088
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.